脳の免疫反応と細胞死

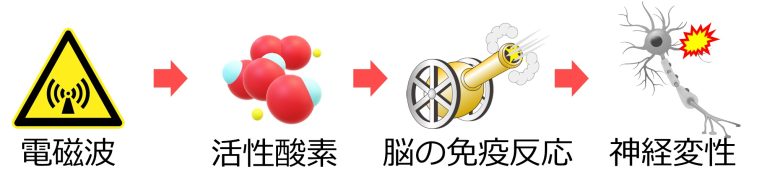
次は、活性酸素が脳の免疫を活性化し、炎症を引き起こすことで神経変性を誘導する経路について解説します。
神経変性とは、不可逆的にニューロンが損傷し、死に至る過程のことです。(Giacalone et al. 2015)
続いて、電磁波によって脳の免疫反応からの神経変性が、実際に起きたを示す研究を紹介します。
目次All_Pages
脳の免疫反応と神経炎症
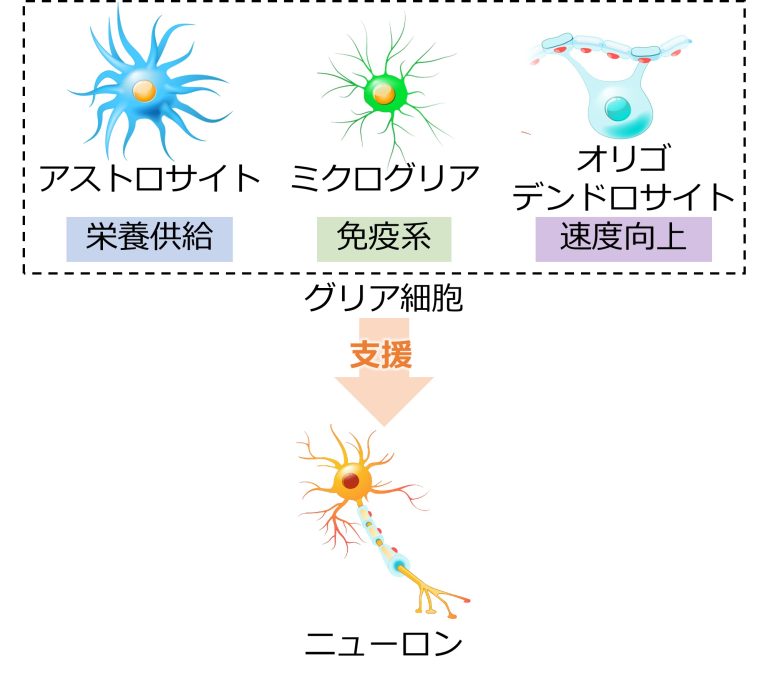
脳における情報処理と伝達を担っているのは、ニューロンという脳細胞です。
脳細胞にはこの他にも、ニューロンに栄養を供給するアストロサイト、脳の免疫系を担うミクログリア、神経伝達速度を高めるオリゴデンドロサイトなどがあり、ニューロンの活動を支えています。
これらの脳細胞はグリア細胞と呼ばれます。
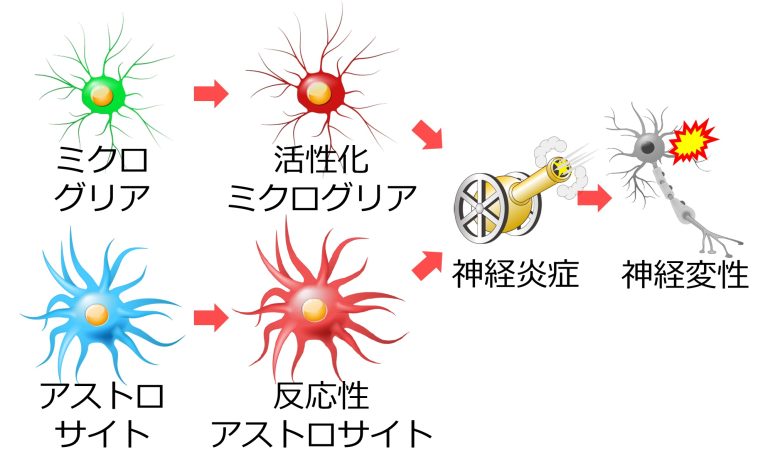
ミクログリアのほか、アストロサイトも脳における免疫反応を担っており、様々な病変に応じて活性化します。
特に活性化したアストロサイトは反応性アストロサイトと呼ばれ、形態も機能も通常のそれとは異なる状態へと変化します。(Ben Haim et al. 2015)
脳における炎症のことを神経炎症といいますが、これらグリア細胞の免疫反応は神経炎症を引き起こし、様々な神経変性疾患に関与していることが知られています。(Heneka et al. 2014)
活性酸素による脳の免疫反応
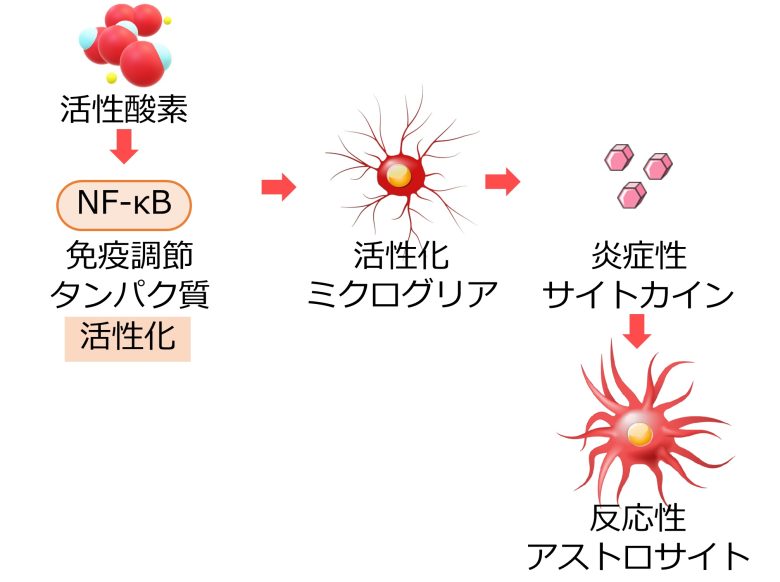
活性酸素は免疫細胞を活性化し、炎症を引き起こすことが知られています。(Chatterjee 2016)
NF-κBは免疫反応の様々な側面を制御するタンパク質ですが (Hayden et al. 2006)、活性酸素はこのNF-κBを活性化することが知られています。(Gloire et al. 2006)
例えば活性酸素はNF-κBの活性化を経由して、免疫細胞であるマクロファージを活性化します。(Rendra et al. 2019)
神経系においても、活性酸素がNF-κBの活性化を経由して、脳内常在のマクロファージである、ミクログリアを活性化します。(Bordt and Polster 2014)
さらに、活性化したミクログリアが、炎症性サイトカインよばれる炎症を促進する物質を周囲に放出し、これをうけてアストロサイトが反応性アストロサイトへと変化することも確認されています。(Liddelow et al. 2017)
このように活性酸素によって活性化したミクログリア、アストロサイトが、神経炎症を引き起こします。
アストロサイトの神経炎症
反応性アストロサイト (活性化アストロサイト) の引き起こす神経炎症は、神経変性を引き起こします。

2015年のフランスの論文が、神経変性疾患における反応性アストロサイトの役割について網羅的に解説されています。(Ben Haim et al. 2015)
この論文をもとに、反応性アストロサイトによる以下の神経変性の経路を紹介します。
- 栄養・抗酸化物質の供給減少
- 興奮毒性への誘導
- グリア伝達物質の放出量の増加
- アミロイドβの大量生成
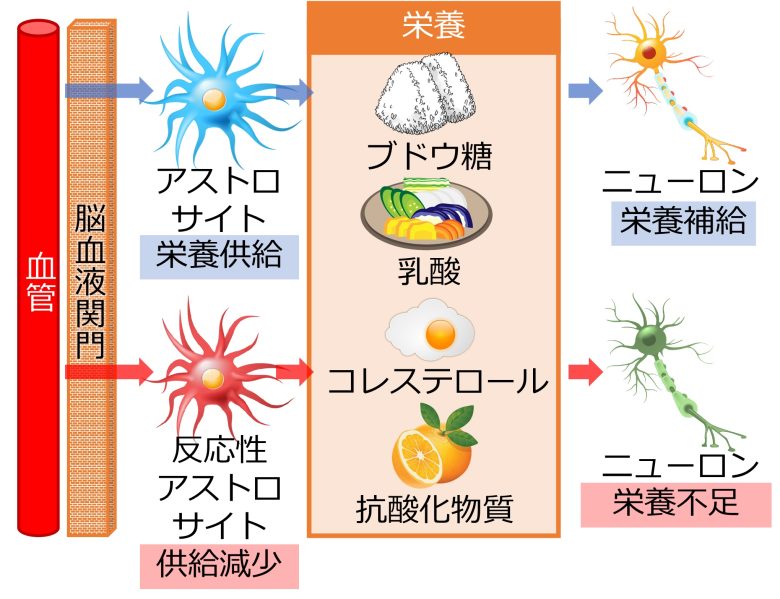
栄養・抗酸化物質の供給減少
脳の血管は脳血液関門によって塞がれているため、ニューロンはアストロサイトから栄養を供給してもらう必要があります。
これらの栄養にはブドウ糖、乳酸、コレステロール、抗酸化物質などがあります。(Ben Haim et al. 2015)
乳酸は脳の重要なエネルギー源で (van Hall et al. 2009)、コレステロールは細胞膜の成分でシナプス形成に必要で、抗酸化物質は酸化ストレスの防御に必要です。
しかしアストロサイトが反応性アストロサイトに変化すると、これらの栄養の供給量が減少していまい (Ben Haim et al. 2015)、ニューロンに有害な作用をもたらしえます。
興奮毒性への誘導
ニューロンの興奮
ニューロンは刺激を受けると活動電位と呼ばれる電気信号を発生させ、これがニューロン内を伝わっていきます。
活動電位が発生している状態のことを「興奮」と呼びます。
活動電位が細胞末端まで到達すると、ニューロンは神経伝達物質と呼ばれる化学信号を放出し、隣接するニューロンを興奮させます。
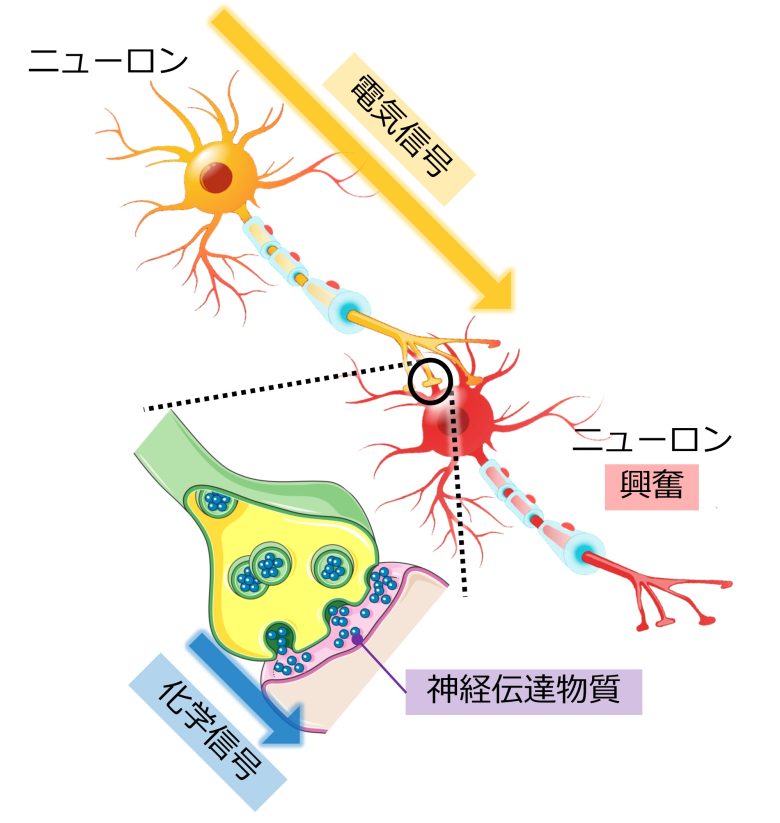
すると隣接するニューロンにおいても活動電位が発生し、細胞末端までそれが伝わると同様に神経伝達物質を放出し、情報が後続のニューロンに伝わっていきます。
このようにして接続された多数のニューロンの連なりが、脳における情報処理と情報伝達の回路を形成しています。
アストロサイトによる興奮毒性
神経伝達物質にはグルタミン酸、ドーパミン、セロトニン、GABAなど、いくつか種類が存在します。
ここでアストロサイトはグルタミン酸を取り込む役割があり、ニューロンが興奮し続けることでニューロンが壊死してしまう、興奮毒性を防いでいます。(Ben Haim et al. 2015)
しかしアストロサイトが反応性アストロサイトへと変化すると、この取り込みが不十分になり、ニューロンは興奮毒性へと陥ります。(Ben Haim et al. 2015)
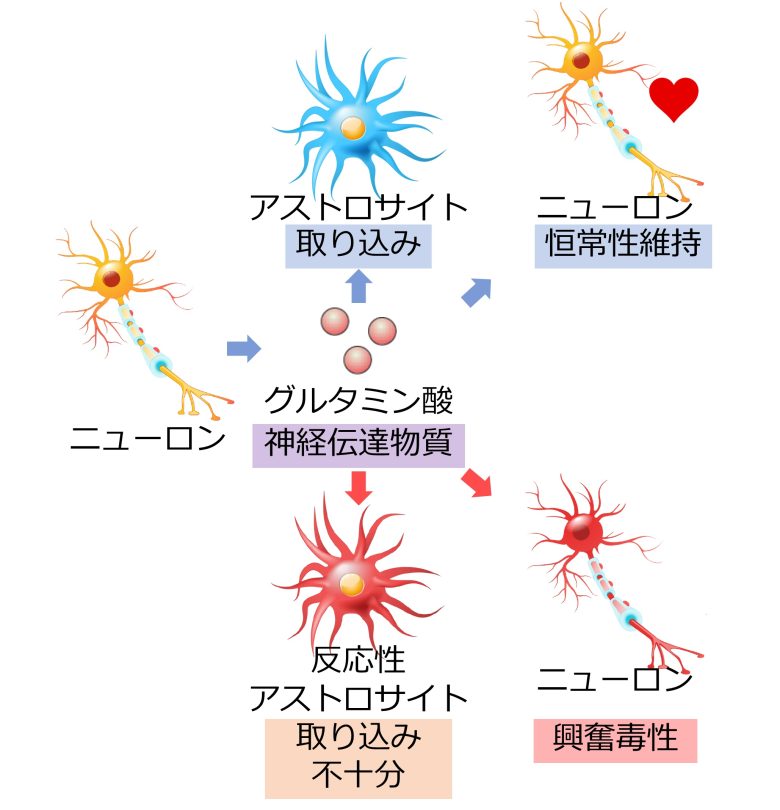
興奮毒性はハンチントン病、アルツハイマー病、ALS、パーキンソン病、多発性硬化症など、多数の神経変性疾患において観察されていることが知られています。(Salińska et al. 2005)
グリア伝達物質の放出量の増加
ニューロンのように、アストロサイト同士が結合して情報伝達を行い、さらに神経伝達物質ならぬグリア伝達物質の放出により、アストロサイトだけでなく、ニューロンを興奮させるという証拠が増えてきています。(Scemes and Giaume 2006)
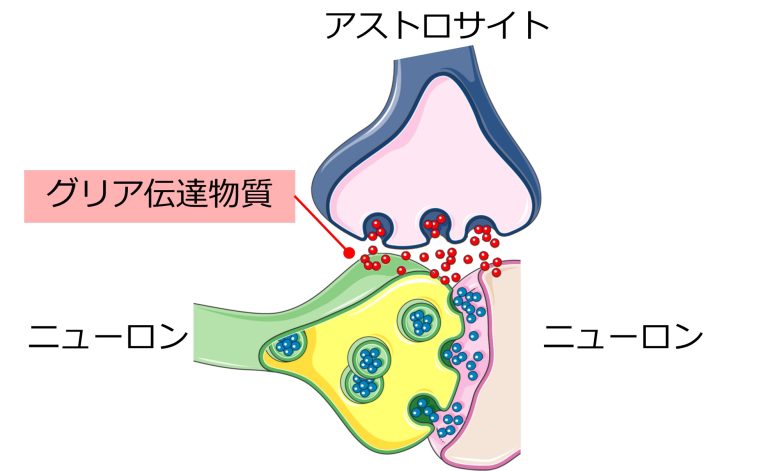
グリア伝達物質としてグルタミン酸、ATP、GABAなどが知られています。(Ben Haim et al. 2015)
ATPは細胞のエネルギー源としてよく知られていますが、意外なことに、グリア伝達物質としても機能し、ニューロンやアストロサイトを活性化します。(Butt 2011)
そしてアストロサイトが反応性アストロサイトへと変化すると、これらグリア伝達物質の放出量が増加し、ニューロンに興奮毒性を引き起こします。(Ben Haim et al. 2015, Ding et al. 2007)
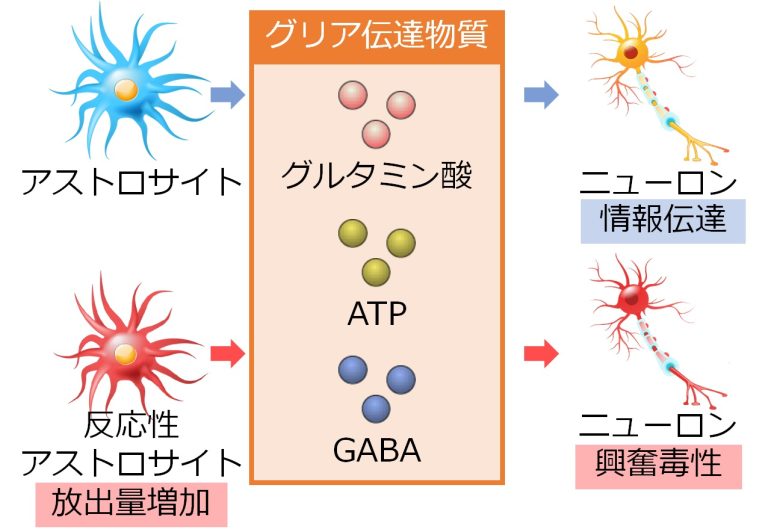
GABAは興奮を抑制するタイプの神経伝達物質ですが、反応性アストロサイトからの放出量が増加することで、マウスにおいて海馬シナプスの成長を抑制し、学習障害と記憶障害を引き起こしたことが確認されています。(Ben Haim et al. 2015, Jo et al. 2014, Wu et al. 2014)
アミロイドβの大量生成
細胞内外のタンパク質の凝集は、神経変性疾患の中心的な特徴です。(Ben Haim et al. 2015)
たとえばアルツハイマー病においては、アミロイドβというタンパク質が細胞外において凝集し、老人斑と呼ばれる沈着物を形成してます。
この老人斑には神経毒性があると考えられています。
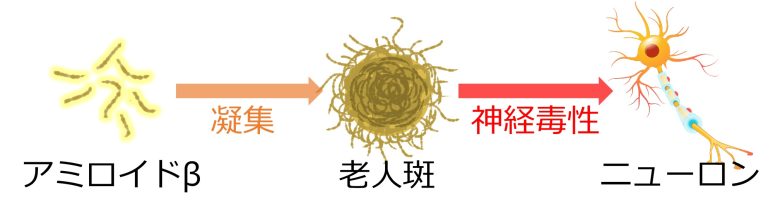
ニューロンのシナプス活動によってこのアミロイドβが生成、細胞外に放出されますが (Cirrito et al. 2005)、アストロサイトによってこれは除去されます(Ben Haim et al. 2015)。
しかし反応性アストロサイトへ変化すると、逆にアミロイドβを大量に生成するようになることが確認されています。(Ben Haim et al. 2015, Zhao et al. 2011)
また、老人斑はアストロサイトを活性化させ、慢性的にアミロイドβを産出させるよう誘導します。(Hu and Van Eldik 1999)
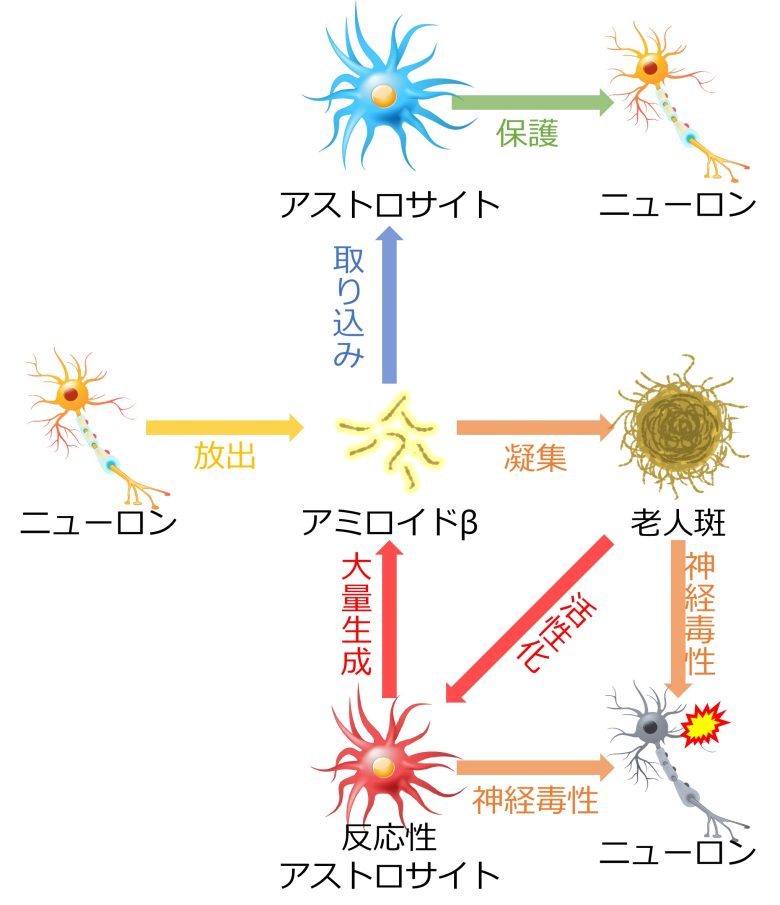
このように慢性的に活性化したアストロサイトは、老人斑とともに神経毒性をもたらすものと考えられます。
ミクログリアの神経炎症
活性化したミクログリアの引き起こす神経炎症も、神経変性を引き起こします。
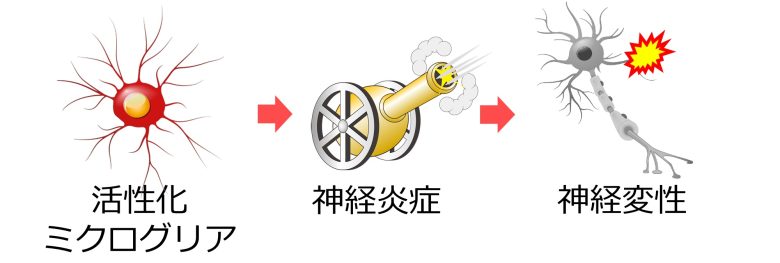
炎症の巻き添え
ミクログリアが活性化すると、細胞膜にNADPHオキシダーゼ、一酸化窒素シンターゼと呼ばれるタンパク質が配備されます。(Brown 2007)
これによりそれぞれ超酸化物、一酸化窒素が生成され、この2つの物質が反応してペルオキシ亜硝酸へと変化します。(Brown 2007)
ペルオキシ亜硝酸には抗ウィルス、抗微生物、抗寄生虫の作用がありますが (Ascenzi et al. 2010)、ニューロンがその巻き添えを被り、細胞死に至ります。(Brown 2007)

また反応性アストロサイトも、このペルオキシ亜硝酸の生成によるニューロン細胞死に関与している可能性があります。 (Ben Haim et al. 2015)
脳の免疫反応による神経変性
以上の話をまとめます。
脳において活性酸素が増加すると、免疫細胞であるミクログリアが活性化し、さらにミクログリアによりアストロサイトが活性化します。
これらの細胞の免疫反応により炎症が引き起こされ、神経変性につながります。
また電磁波は脳において活性酸素を増加させることが確認されています。
したがって電磁波は脳において免疫反応を発生させて炎症を引き起こし、最終的に神経変性をもたらす、という結論が導き出せます。
電磁波による脳の免疫反応と神経変性
ここで電磁波被曝で脳の免疫反応が発生し、さらに神経変性が起きたことを示す研究を紹介します。
研究紹介
Yang et al. 2010
マウスのミクログリアが、パルス変調した2.45 GHzの高周波電磁波を培地平均SAR 6 W/kgで20分間だけ被曝しました。
するとミクログリアが活性化し、アスロトサイトを活性化させるシグナルである炎症性サイトカインの放出量が増加しました。
ミクログリアの活性化
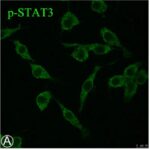
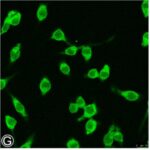
電磁波被曝でミクログリア活性化の指標である、STAT3のリン酸化が増加しました。
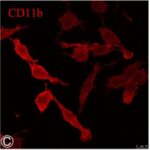
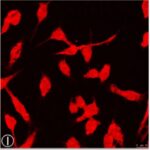
電磁波被曝でミクログリア活性化の指標である、CD11bの発現量が増加しました。
炎症性サイトカインの
放出量の増加
電磁波被曝からの経過時間が長くなるほど、炎症性サイトカインであるTNF-αの放出量が増加しました。
Megha et al. 2015
オスラットが900MHz、1800MHz、2450MHzの高周波電磁波をそれぞれ全身平均SAR 0.00059 W/kg、0.00058 W/kg、0.00066 W/kgで1日2時間、60日間被曝しました。
するとラットの海馬において、電磁波の周波数が高くなるほど、活性酸素が増加し、アスロトサイトを活性化させるシグナルである、炎症性サイトカインの放出量が増加しました。
活性酸素の増加
脂質過酸化の増加と抗酸化活性の減少は活性酸素の増加を意味します。
炎症性サイトカインの増加
MAUSSETBONNEFONT et al. 2004
成体のオスラットが、携帯電話の電磁波に似せた高周波電磁波を脳平均SAR 6 W/kgで15分間だけ被曝しました。
するとラットの脳において、大脳皮質、海馬、そして特に大脳基底核の線条体で反応性アストロサイトが増加しました。
反応性アストロサイトの増加 1
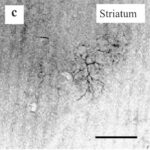
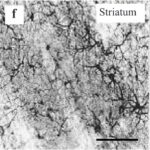
わずか15分間の電磁波被曝で、大脳基底核の線条体において反応性アストロサイトが増加しました。
反応性アストロサイトの増加 2
反応性アストロサイトの指標が、わずか15分間の電磁波被曝で大脳皮質で25%、海馬で20%、そして大脳基底核の線条体で140%増加しました。
Ammari et al. 2010
子どもに相当する生後6週のオスラットに、携帯電話の電磁波に似せた高周波電磁波を、脳平均SAR 1.5 W/kgで1日45分、あるいは6 W/kgで1日15分、8週間被曝させました。
すると被曝から3日後のラットの脳において、前頭前野、海馬の歯状回、大脳基底核の線条体と淡蒼球、小脳皮質で、反応性アストロサイトが増加しました。
反応性アストロサイトの増加
Akakin et al. 2020
局所SAR 1.79 W/kgの携帯電話を飼育ケージの上に設置して、1日2時間待機中か会話中にし、妊娠中のラットがその電磁波を妊娠初期の1週間被曝、さらに生まれた仔ラットが2ヶ月間被曝しました (乳児・子ども期に相当)
すると仔ラットの脳において、携帯電話の出力が強いほど、また被曝期間が長いほど、反応性アストロサイトが増加しました。
さらに大脳皮質、海馬歯状回、海馬アンモン角でニューロンが変性し、また三叉神経とその髄鞘が変性しました。
反応性アストロサイトの増加
神経変性
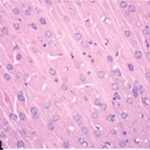
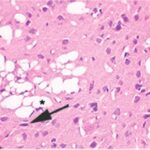
胎児・乳児・子ども期の携帯電話の電磁波被曝で大脳皮質に無数の空胞が発生し、ダークニューロンとみられる、縮んで暗く染まった、変性したニューロンが多数出現しました。
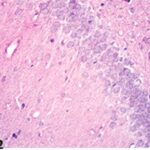
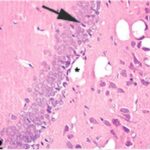
胎児・乳児・子ども期の携帯電話の電磁波被曝で海馬歯状回に無数の空胞が発生し、ダークニューロンとみられる、縮んで暗く染まった、変性したニューロンが多数出現しました。
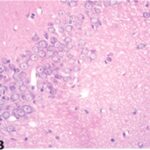
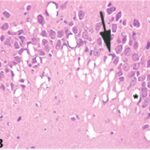
胎児・乳児・子ども期の携帯電話の電磁波被曝で海馬アンモン角に無数の空胞が発生し、ダークニューロンとみられる、縮んで暗く染まった、変性したニューロンが多数出現しました。
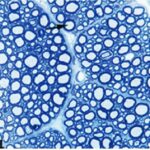
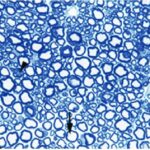
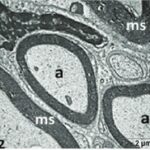
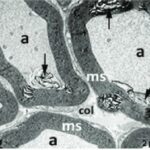
胎児・乳児・子ども期の携帯電話の電磁波被曝で三叉神経の軸索とその髄鞘が変性しました。
Afeefy et al. 2013
局所SAR 0.43 W/kgの携帯電話を飼育ケージの外側に設置して、1日2時間着信中にし、妊娠中のラットがその電磁波を妊娠全期の3週間被曝し、さらに生まれた仔ラットが4週間被曝しました(乳児期に相当)。
すると仔ラットの海馬において、反応性アストロサイトが増加しました。
さらに歯状回とアンモン角でニューロンが変性しました。
反応性アストロサイトの増加
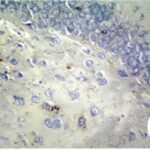
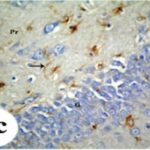
胎児期と幼児期の携帯電話の電磁波被曝で、海馬歯状回で反応性アストロサイトが増加しました。
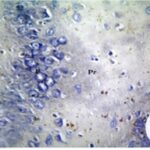
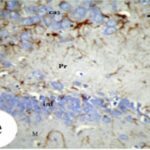
胎児期と幼児期の携帯電話の電磁波被曝で、海馬アンモン角で反応性アストロサイトが増加しました。
海馬歯状回で神経変性
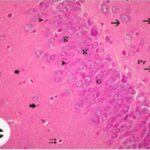
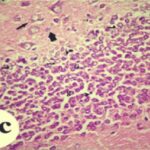
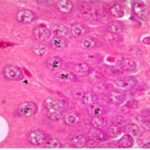
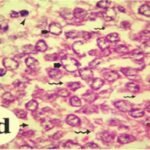
胎児・乳児期の携帯電話の電磁波被曝で、海馬歯状回でニューロン (顆粒細胞) が変性しました。
海馬アンモン角で神経変性
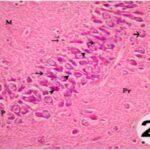
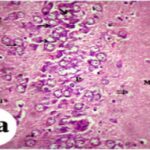
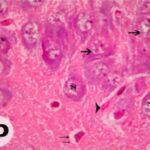
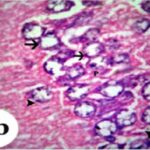
胎児・乳児期の携帯電話の電磁波被曝で、海馬アンモン角でニューロン (錐体細胞) が変性しました。
続いて同様の研究から成体のラットを対象にした実験です。
局所SAR 0.43 W/kgの携帯電話を飼育ケージの外側に設置して、1日2時間着信中にし、成体のラットがその電磁波を12週間被曝しました。
すると成体ラットの海馬において、反応性アストロサイトが増加しました。
さらに歯状回とアンモン角でニューロンが変性しました。
反応性アストロサイトの増加
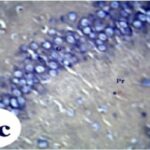
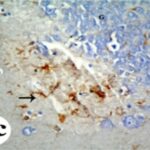
携帯電話の電磁波被曝で、海馬歯状回で反応性アストロサイトが増加しました。
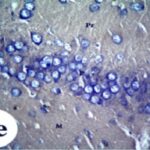
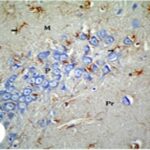
携帯電話の電磁波被曝で、海馬アンモン角で反応性アストロサイトが増加しました。
海馬歯状回で神経変性
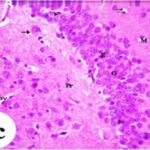
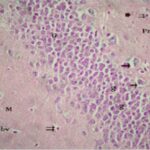
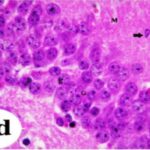
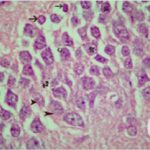
携帯電話の電磁波被曝で、海馬歯状回でニューロン (顆粒細胞) が変性しました。
海馬アンモン角で神経変性
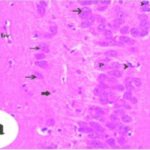
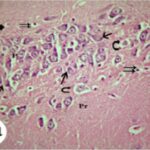
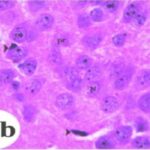
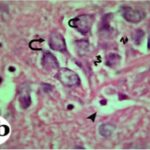
携帯電話
携帯電話の電磁波被曝で、海馬アンモン角でニューロン (錐体細胞) が変性しました。
実験の紹介は以上です。
実際に電磁波は脳に免疫反応を引き起こし、神経変性をもたらすことを確認しました。


