複製ストレス

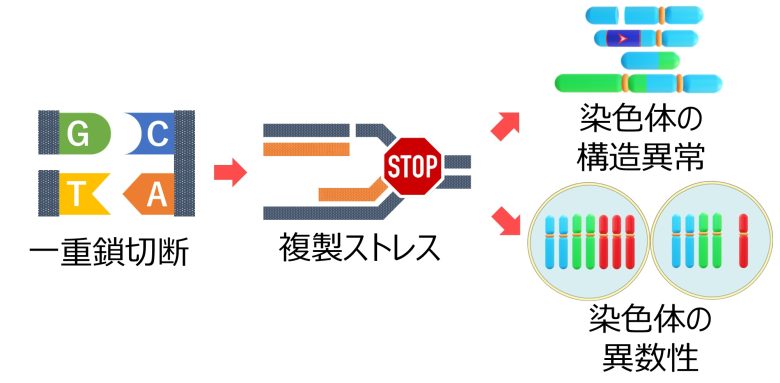
一重鎖切断から複製ストレスが発生すると、さらに大規模な突然変異である染色体の構造異常や異数性が引き起こされます。
主に2015年のフランスの複製ストレスに関する総説をもとに、その仕組みを説明します。(Gelot et al. 2015)
続いて、ここまで説明してきた各種の突然変異が、実際に電磁波によって引き起こされたことを示す研究を紹介します。
ある実験では携帯電話の電磁波の9時間の被曝と、放射能20分の被曝が同程度の染色体異常を引き起こすことを示しており、電磁波がいかに有害か物語ります。
染色体の構造異常
まず複製ストレスがどのように染色体の構造異常を引き起こすのか、説明します。

二重鎖切断の形成
DNA複製中に一重鎖切断に遭遇すると、DNA複製はそこで停止し、複製ストレスが発生します。
ここでの大きな問題は、元のDNAの一重鎖切断が、新しいDNAとの間で二重鎖切断を形成してしまうことです。(Gelot et al. 2015)
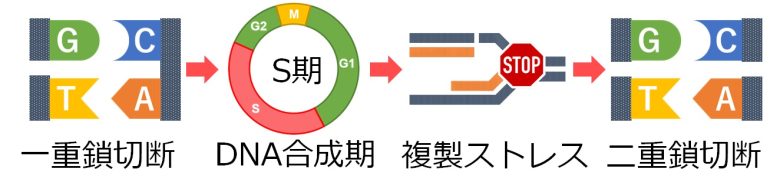
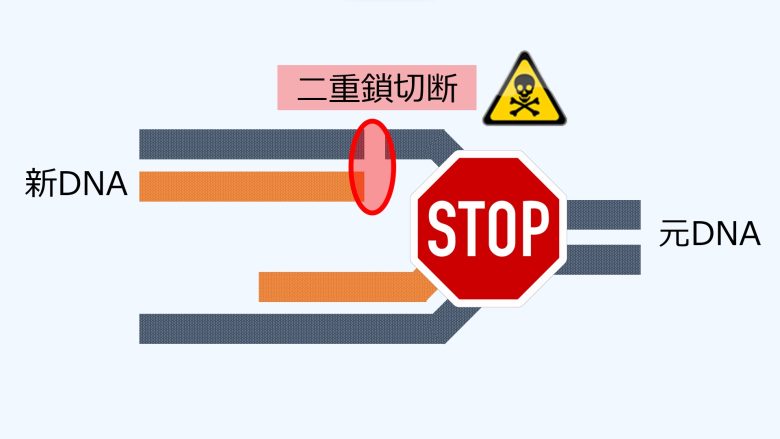
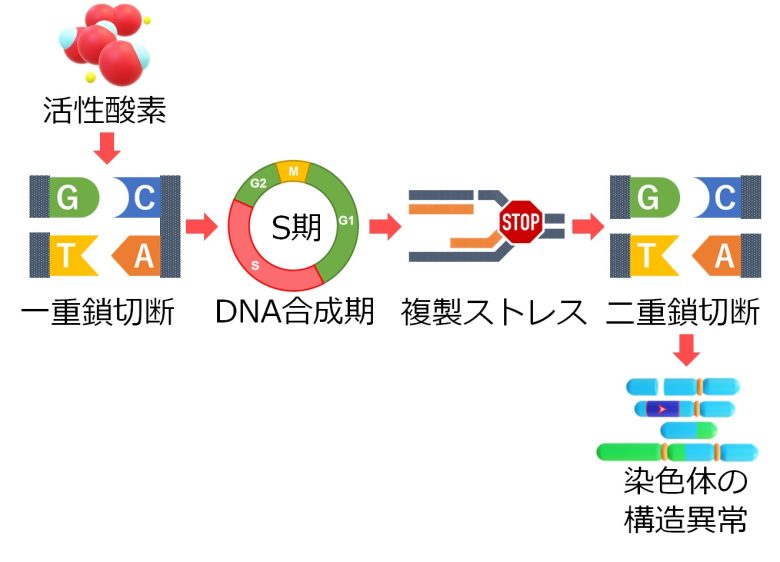
つまり、活性酸素を増加させるとDNAの一重鎖切断が増加しますが、これがそのままDNA合成期における二重鎖切断の増加につながるということです。
従って二重鎖切断の修正ミスも増加することになり、結果として染色体の欠失や逆位、転座、無動原体、二動原体といった、染色体の構造異常が増加してしまうことになります。
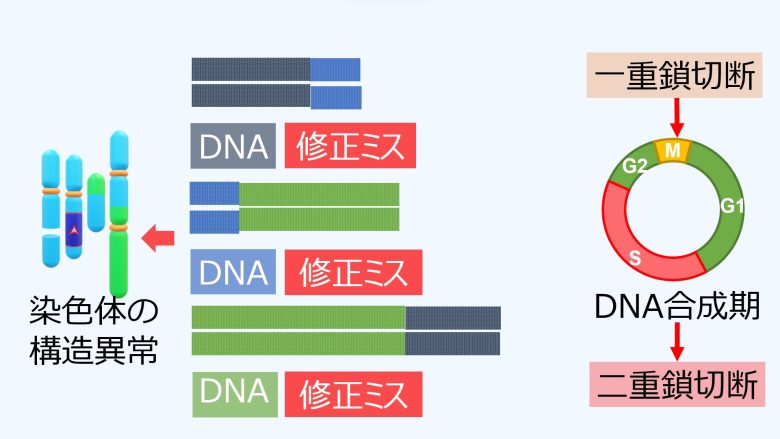
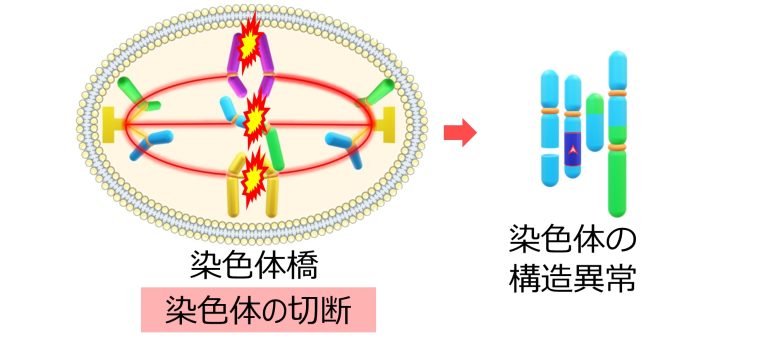
染色体橋の形成
複製ストレスにより一重鎖切断は二重鎖切断へと変換され、二重鎖切断の修復ミスによって様々な染色体の構造異常が生まれます。
このなかでも二動原体は、動原体を2つもつことから、染色体橋を形成し、細胞分裂を停止させます。
この他にも複製ストレスにより、DNAが未複製のままになり、あるいは複製したDNAが絡み合い、これによっても染色体橋が形成されます。(Gelot et al. 2015)
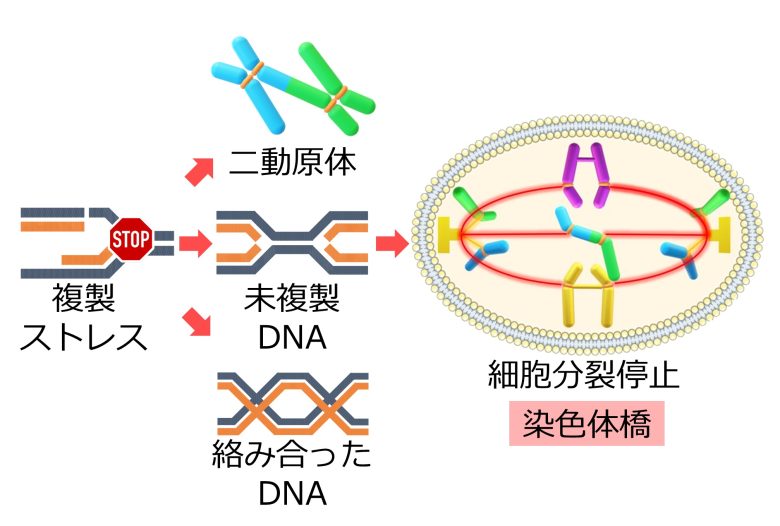
これらの染色体橋は染色体の切断をもって解消されるため、染色体の構造異常を悪化させてしまいます。(Gelot et al. 2015)
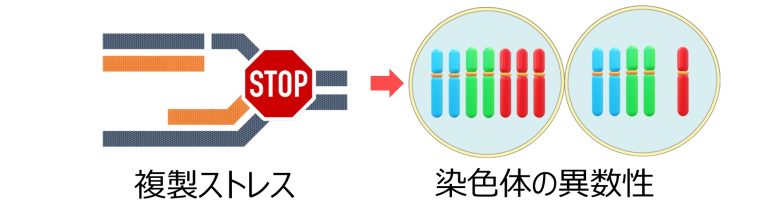
染色体の異数性
次に複製ストレスがどのように染色体の異数性を引き起こすのか、説明します
染色体の異数性とは?
ヒト染色体は父親と母親から受け続いだ1本ずつの染色体が2本で対になり、合計23対、46本存在します。うち1対は性別を決定するXとYの性染色体です。

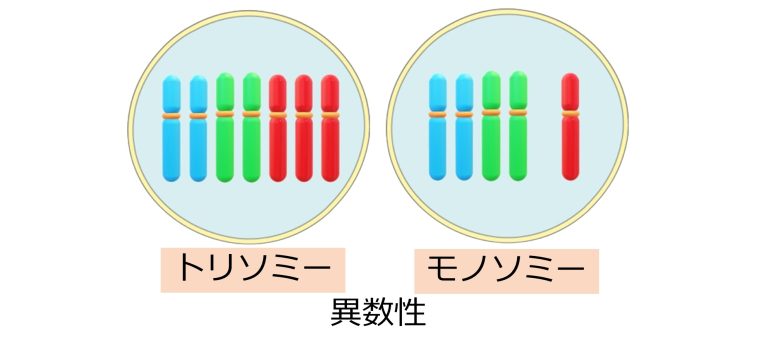
そして染色体の異数性とは、この染色体の数が多かったり少なかったりすることを言います。
特に1対の染色体の数 (=2本) が3本に増えていることをトリソミーといい、1本に減っていることをモノソミーといいます。
複製ストレスからの異数性
ここで2019年のスイスの研究をもとに、複製ストレスが染色体の異数性を引き起こす経路を紹介します。(Wilhelm et al. 2019)
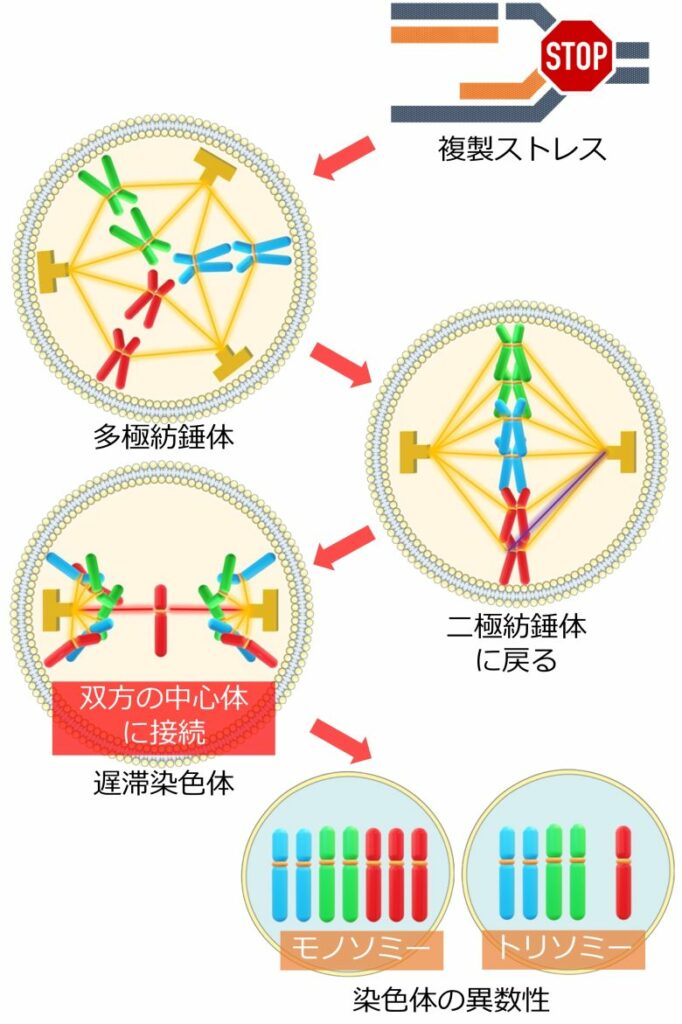
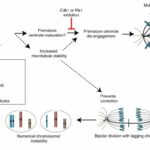
複製ストレスから異数性の経路
(Wilhelm et al. 2019から引用・改変)
- 複製ストレスによって余剰な中心体が出現し、多極紡錘体 (中心体が3つ以上) が形成される。
- 多極紡錘体は通常、二極紡錘体に戻される。
- しかしその修正の過程で、遅滞染色体と呼ばれる、紡錘体の両極に接続されてしまった染色体が出現する。
- 遅滞染色体の誤った分配により、別々であるべき染色体が同一の細胞核に属してしまう。
- これにより片方の細胞では染色体が1本多くなり (トリソミー)、もう片方は1本少なくなる (モノソミー)、染色体の異数性が発生する。
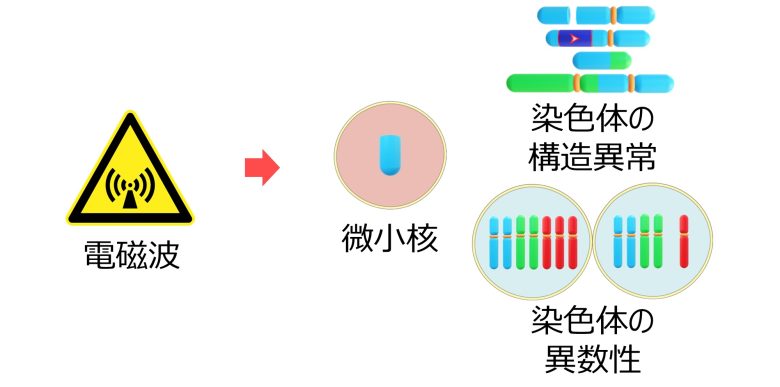
また遅滞染色体は、どちらの細胞核に属することができずに、微小核を形成することもあります。これによっても染色体の異数性、特にモノソミーが発生します。(Norppa 2003)
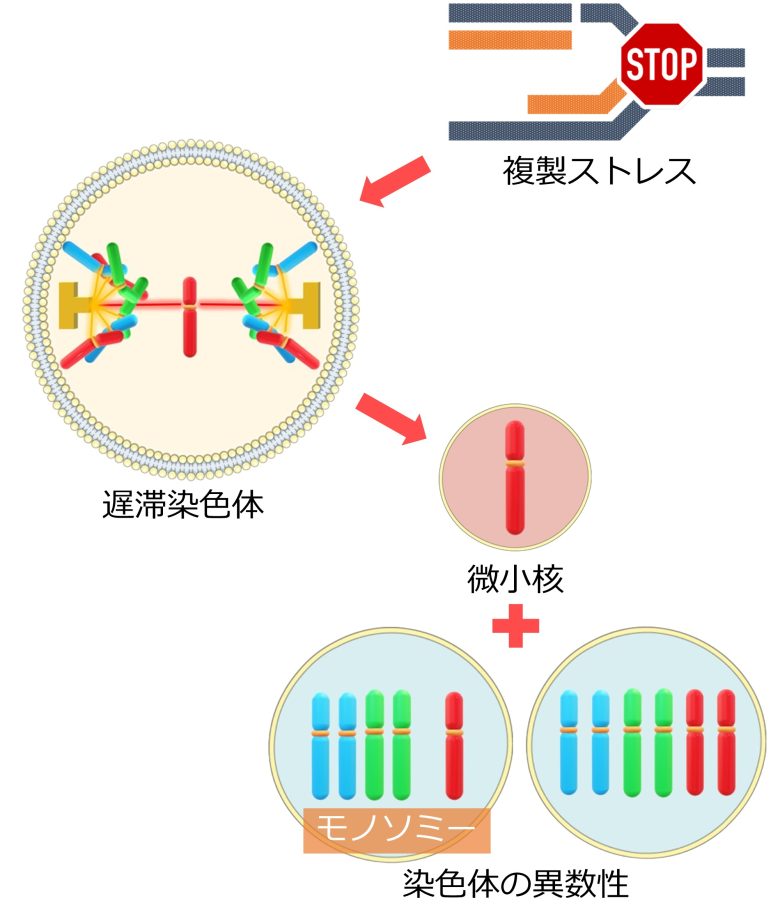
哺乳類細胞において、紡錘体の両極から接続された染色体、つまり遅滞染色体は、異数性の主要な機構です。 (Cimini et al. 2001)
複製ストレスのまとめ
以下は複製ストレスから染色体異常の経路のまとめです。
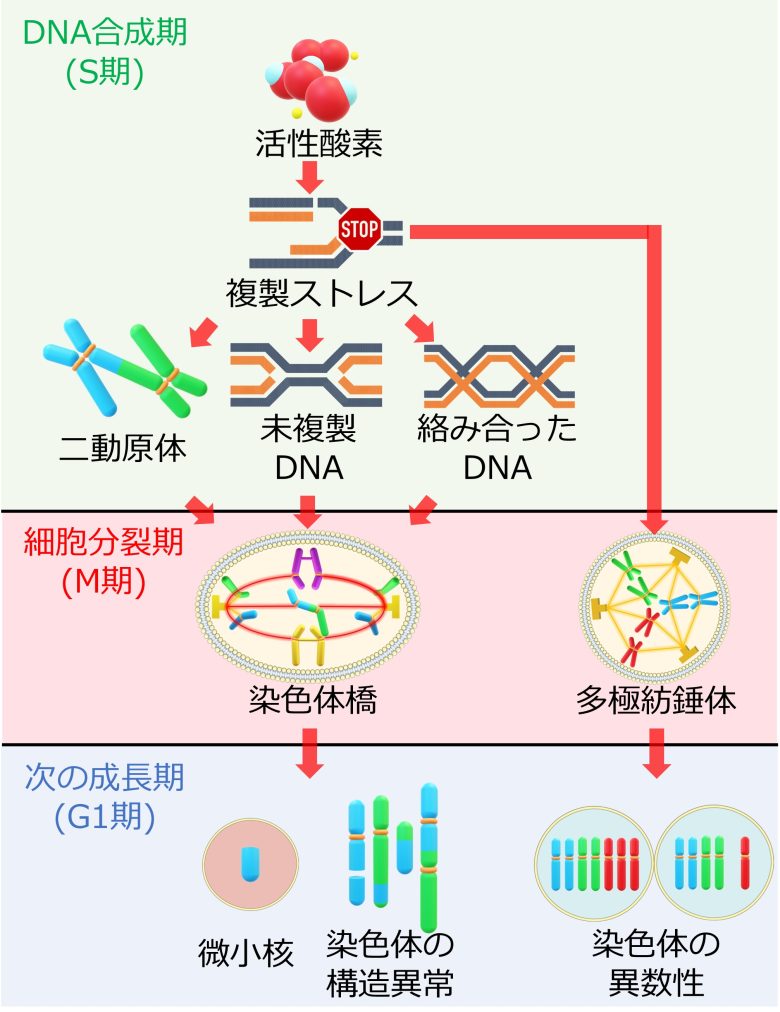
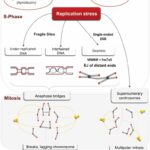
複製ストレスから染色体異常の経路
(Gelot et al. 2015から引用・改変)
- 活性酸素などの要因によって、DNA合成期において複製ストレスが発生する。
- 複製ストレスは二動原体などの染色体の構造異常を引き起こす。
- またDNAを未複製のままにし、また複製したDNAを絡み合わせる。
- これらの異常が細胞分裂期まで至ると細胞分裂は停止し、染色体橋が形成される。
- 染色体橋が染色体の切断によって解消され、染色体の構造異常が悪化する。
- また複製ストレスにより多極紡錘体が形成される。
- 多極紡錘体は染色体の誤分配につながり、染色体の異数性が発生する。
突然変異のまとめ
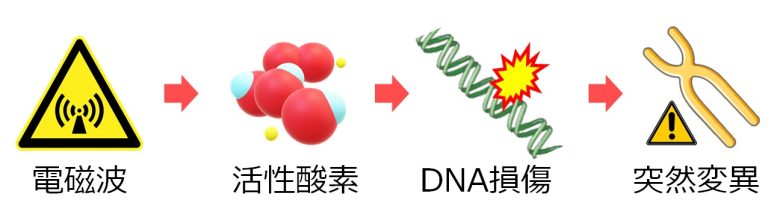
ここまでの、複製ストレス含めた、DNA損傷からの突然変異の経路を以下の図にまとめました。
電磁波による突然変異
ここで電磁波被曝により突然変異が起きたことを示す研究を紹介します。
研究紹介
Pesnya and Romanovsky 2013
局所SAR 1.4 W/kgの携帯電話を設置し、1日1時間または3時間会話中にし、発芽したての玉ねぎの球根が1.5 cmの距離でその電磁波を3日間被曝しました。
すると被曝時間が長くなるほど、染色体橋、遅滞染色体、chromosome stickiness、染色体の断片、微小核が増加しました。
また、放射能 (プルトニウム239のアルファ粒子) を被曝したグループと比較したところ、9時間 (1日3時間×3日) の携帯電話の電磁波被曝による損傷は、20分間の放射能被曝に匹敵するものでした。
正常な細胞分裂
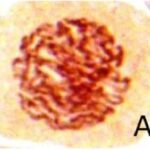

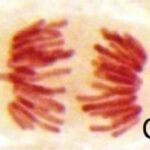
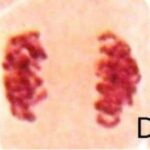
対照群では正常な細胞分裂が観察されました。
染色体橋の増加 1
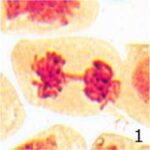
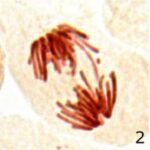
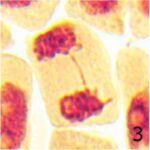
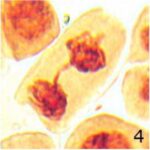
9時間の携帯電話の電磁波被曝で染色体橋が増加しました。
染色体橋の増加 2
染色体橋の頻度が、9時間の携帯電話の電磁波被曝で9倍になり、20分の放射能被曝で4倍になりました。
遅滞染色体の増加 1
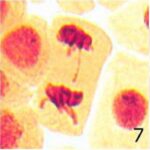
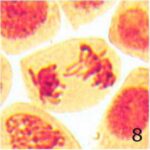
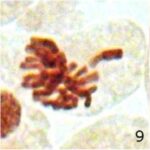
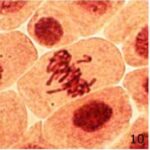
9時間の携帯電話の電磁波被曝で遅滞染色体が増加しました。
遅滞染色体の増加 2
遅滞染色体の頻度が、9時間の携帯電話の電磁波被曝で18倍になり、20分の放射能被曝で3倍になりました。
chromosome stickinessの増加 1
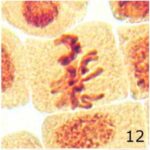
9時間の携帯電話の電磁波被曝でchromosome stickinessが増加しました。
chromosome stickinessの増加 2
chromosome stickinessの頻度が、9時間の携帯電話の電磁波被曝で20倍になり、20分の放射能被曝で13倍になりました。
染色体の断片の増加 1
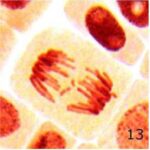
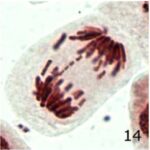
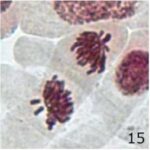
9時間の携帯電話の電磁波被曝で染色体の断片が増加しました。
染色体の断片の増加 2
染色体の断片の頻度が、9時間の携帯電話の電磁波被曝で5倍になり、20分の放射能被曝で16倍になりました。
微小核の増加 1
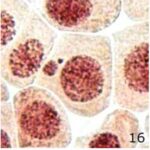
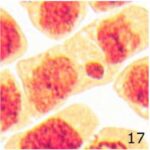
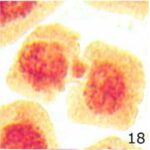
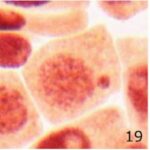
9時間の携帯電話の電磁波被曝で微小核が増加しました。
微小核の増加 2
微小核の頻度が、9時間の携帯電話の電磁波被曝で14倍になり、20分の放射能被曝で23倍になりました。
その他の異常
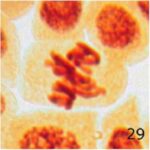
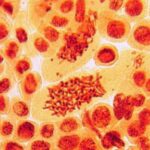
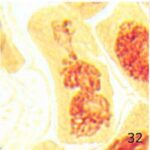
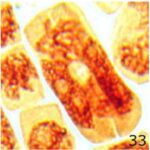
携帯電話の9時間の電磁波被曝で多極細胞分裂、巨大な倍数体細胞、特定不能の異常が発生しました。
D’Ambroslo et al. 1985
ウシから血液を採取し、血中リンパ球が強さ16 mV/mで50Hzの低周波電磁波を72時間被曝しました。
すると染色体の切断、構造異常、異数性、断片、倍数性が増加しました。
染色体の切断の増加
染色体の切断の割合が、電磁波被曝で3倍になりました。
染色体の構造異常の増加
染色体の構造異常の割合が、電磁波被曝で10倍になりました。
染色体の異数性の増加
染色体の異数性の割合が、電磁波被曝で3倍になりました。
染色体の断片の増加
染色体の断片の割合が、電磁波被曝で2倍になりました。
染色体の倍数性の増加
染色体の倍数性の割合が、電磁波被曝で5倍になりました。
Zothansiama et al. 2017
インドのミゾラム州アイザウルの研究参加者において、自宅から携帯基地局までの距離が短くなるほど、血液中の活性酸素が増加し、血中リンパ球で微小核が増加しました。
高周波電磁波の強さは、携帯基地局の300m圏外で平均 0.0035 μW/cm2、80m圏内で平均 0.5 μW/cm2でした。
活性酸素の増加
脂質過酸化の増加、抗酸化活性の減少は活性酸素の増加を意味します。
微小核の増加
微小核の頻度が、携帯基地局の20m圏内で、7割増加しました。
Mairs et al. 2007
ヒト神経膠腫細胞が、強さ1mTで50Hzの低周波電磁波を12時間被曝しました。
するとDNAの突然変異が増加しました。主な突然変異はヘテロ接合性喪失とアレル不均衡で、双方ともに単一遺伝子の喪失が増加したことを意味します。
また、0.3あるいは3Gyの電離放射線と低周波電磁波を組み合わせると、電離放射線による突然変異が増幅しました。(※)
原因として、電磁波が細胞周期チェックポイントを弱めている可能性があります (6ページ目参照)。
突然変異の増加
Othman et al. 2001
エジプトの男性航空管制官と技師、ならびに無作為に選出した男性について、レーダースクリーン・アンテナ・衛星施設・閉回路テレビ (CCTV) からの電磁波の職業被曝を受けていると、血中リンパ球で7番・17番・Y染色体の異数性が増加しました。
異数性の増加
異数性 (モノソミー、喪失) の割合が、レーダースクリーン・アンテナ・衛星施設・CCTVからの電磁波の職業被曝で、7倍染色体で6倍、17番染色体で4倍、Y染色体で4倍になりました。
Mashevich et al. 2003
イスラエルの男性の研究参加者から血液を採取し、血中リンパ球が830MHzの高周波電磁波を培地平均SAR 2-8.2 W/kgで72時間被曝しました。
すると17番染色体において、SAR値が大きくなるほど、異常なDNA複製と異数性が増加しました。
異常なDNA複製の増加
17番染色体において、SAR値が大きくなるほど、異常なDNA複製の割合が増加しました。
異数性の増加
17番染色体において、SAR値が大きくなるほど、異数性の割合が増加しました。
Manikowska-Czerska et al. 1985
青年に相当する生後8-10週のマウスが、2.45 GHzの高周波電磁波を全身平均SAR 0.05-20 mW/kgで1日30分、2週間被曝しました。
すると精巣内の精子形成細胞において、染色体の構造異常である転座と、一価染色体が増加しました。
一価染色体とは、細胞分裂時に対になってない染色体のことで、遅滞染色体と同様、異数性を引き起こします。(Nagaoka et al. 2012)
転座の増加
転座の割合が、高周波電磁波の被曝で増加しました。
一価染色体の増加
一価染色体の割合が、高周波電磁波の被曝で増加しました。
Wi-Fiと同じ周波数の電磁波が、全身平均SAR0.00005W/kgという微弱な強さでも生体効果を示したことは、特筆すべき点といえます。
Balamuralikrishnan et al. 2012
インドのタミル・ナードゥ州コインバトールの研究参加者について、高圧変電所の内部・近隣で働いていると、血中リンパ球で染色体切断や微小核が増加しました。
染色体切断の増加
染色体切断の頻度が、高圧変電所の近隣での勤務で2倍、内部での勤務で3倍になりました。
微小核の増加
微小核の頻度が、高圧変電所の近隣での勤務で3倍、内部での勤務で3倍になりました。
実験の紹介は以上です。
実際に電磁波は微小核、染色体の構造異常、異数性といった突然変異を引き起こすことを確認しました。